


This content is archived from the Feline Nutrition Foundation
The Skinny on Senior Cats: Metabolism Explained
- Updated: Sunday, April 07, 2019 02:34 PM
- Published: Saturday, March 19, 2016 09:21 PM
- Written by Mark E. Peterson, DVM, Dip. ACVIM
 In this article, my mission is to review energy and protein metabolism in cats. I will attempt to explain why, unfortunately, increasing the amount of fat or carbohydrate fed to an older or hyperthyroid cat generally cannot compensate for a diet deficient in an optimal protein content. As I discussed in in my previous article, energy requirements sharply and progressively increase in older cats starting at ten to twelve years of age.¹⁻³ If daily caloric intake is not increased, progressive weight loss will result, due in large part to the loss of lean body mass, i.e., muscle mass, a phenomenon referred to as sarcopenia of aging.⁴⁻⁷ In addition to this increased caloric intake, older cats also require higher amounts of protein to maintain protein reserves compared with younger adult cats.³ ⁸⁻¹¹ As cats age, they absorb and metabolize protein less efficiently.¹⁰ Therefore, it's extremely important to feed high-quality protein, i.e., animal source rather than grain-based, as well as an adequate quantity of protein to aging cats.
In this article, my mission is to review energy and protein metabolism in cats. I will attempt to explain why, unfortunately, increasing the amount of fat or carbohydrate fed to an older or hyperthyroid cat generally cannot compensate for a diet deficient in an optimal protein content. As I discussed in in my previous article, energy requirements sharply and progressively increase in older cats starting at ten to twelve years of age.¹⁻³ If daily caloric intake is not increased, progressive weight loss will result, due in large part to the loss of lean body mass, i.e., muscle mass, a phenomenon referred to as sarcopenia of aging.⁴⁻⁷ In addition to this increased caloric intake, older cats also require higher amounts of protein to maintain protein reserves compared with younger adult cats.³ ⁸⁻¹¹ As cats age, they absorb and metabolize protein less efficiently.¹⁰ Therefore, it's extremely important to feed high-quality protein, i.e., animal source rather than grain-based, as well as an adequate quantity of protein to aging cats.
Animals derive energy from the oxidation of the macronutrients carbohydrate, fat and protein.¹² Most animals, including rats and humans, generally adapt to the diet being fed and can oxidize whatever fuel mixture is contained in their prevailing diet.¹³ ¹⁴ This enables most animals, especially omnivores, to use widely differing diet compositions to satisfy their energy requirements.
A number of veterinarians have contacted me with concerns about the potential problems associated with feeding a high protein diet to the geriatric cat, especially in those cats with kidney disease. Basically, the questions come down to the following:
- If animals can adapt their energy intake based on a wide variety of feeding regimens, why can't we just increase the amounts of fat and carbohydrate fed to compensate for the increasing energy requirements in our senior and hyperthyroid cats? Won't that help prevent loss of muscle mass?
- Can cats as obligate carnivores adapt to the same extent?
Overview of the Body's Energy Production - Nutritional Biochemistry 101
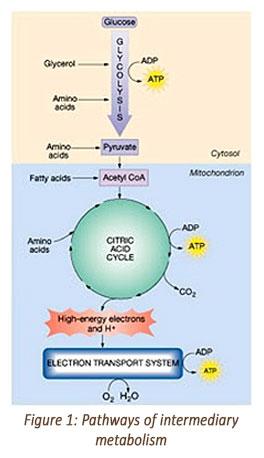
Let's start with a brief review of nutritional biochemistry and the metabolic pathways an animal uses for energy production. The initial biochemical reactions by which energy is derived from carbohydrates, fats and proteins are different. However, all three macronutrients eventually go through a final common pathway for energy generation called the citric acid cycle - also known as the tricarboxylic acid cycle, the TCA cycle or the Krebs cycle.¹² ¹⁵ Please see Figure 1.
Carbohydrates: Glucose derived from dietary carbohydrates is first oxidized through the glycolysis pathways to yield pyruvate and then acetyl-CoA. The acetyl-CoA is then oxidized in the citric acid cycle.¹² ¹⁵ High-energy electrons produced in the citric acid cycle enter the electron transport chain to generate adenosine triphosphate, ATP, which transfers its energy from chemical bonds to energy-absorbing chemical reactions within the cell.
Fats: Fatty acids from dietary fats are initially oxidized to acetyl-CoA by the beta-oxidation pathway, which then enter the citric acid cycle and subsequently generate ATP via oxidative phosphorylation in the electron transport system.
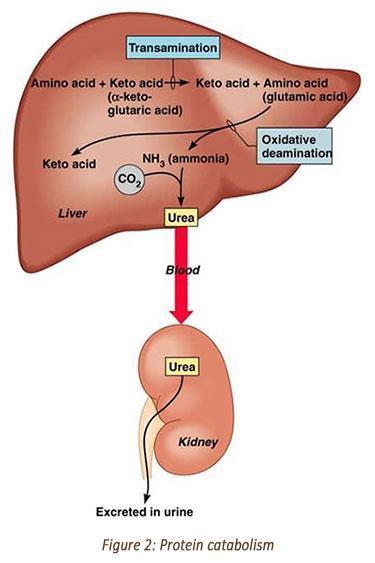
 Protein: Once protein synthesis is maximized, excess amino acids can be used for energy by undergoing transamination and deamination. In oxidative deamination, an amino group is removed from the amino acid and converted to ammonia.
Protein: Once protein synthesis is maximized, excess amino acids can be used for energy by undergoing transamination and deamination. In oxidative deamination, an amino group is removed from the amino acid and converted to ammonia.
The remaining carbon skeleton from these deaminated amino acids, organic ketoacids, can be recycled to make nonessential amino acids, or they can be oxidized for energy.¹² ¹⁵ If used for energy, the catabolized amino acids enter either the glycolysis pathway or the citric acid cycle to ultimately form ATP in the electron transport system. The carbon skeletons of catabolized amino acids can also be converted to glucose or ketone bodies in the liver.
Ammonia is toxic to the body, so enzymes convert it to urea by addition of carbon dioxide in the urea cycle, which takes place in the liver. Urea can safely diffuse into the blood and then be excreted into the urine. Please see Figure 2.
Interconversion of nutrient molecules: In metabolism, there commonly is interconversion of nutrient molecules. Excess glucose is stored as glycogen, this is known as glycogenesis; when glycogen stores are filled, glucose and amino acids are used to synthesize lipids, this is lipogenesis. Therefore, glycogen and adipose tissue both serve as long-term forms of energy storage in the body.
In contrast, amino acids are not "stored" in the body, other than that found in muscle and other structural proteins. Although amino acids can be converted to either fat or glucose, the opposite does not occur - fat and carbohydrates cannot be directly converted to amino acids to be made into protein.¹² ¹⁵
Protein Structure, Functions & Metabolism
Proteins, from the Greek proteios meaning "first," are important biological molecules that consist of strings of amino acids linked together in sequence as polypeptide chains.16 Proteins vary in shape and size, some consisting of only 20-30 amino acids and others of several thousands.¹² ¹⁵
Proteins are present in every living cell. In the skin, hair, cartilage, muscles, tendons and ligaments, proteins hold together, protect and provide structure to the body. As enzymes, hormones, antibodies and globulins, they catalyze and regulate body chemistry. In addition, protein is required for growth and tissue repair, as well as for maintaining muscle mass and tone.
 Essential and Nonessential Amino Acids
Essential and Nonessential Amino Acids
More than 20 amino acids are involved in the synthesis of protein in the body. Essential amino acids are those that cannot be formed in sufficient amounts to meet the requirements for growth and maintenance and must be supplied in the diet. Nonessential amino acids are those that the body can produce in sufficient amounts from other nutrients and metabolites and thus do not need to be supplied in the diet.
Even with the nonessential amino acids, however, fat and carbohydrates cannot be directly converted to amino acids to be made into protein.¹² ¹⁵ Although all mammals can synthesize ten nonessential amino acids from precursor carbon skeletons, if adequate nitrogen and energy are available, there is no direct way that amino acids can be synthesized directly from either fat or carbohydrates.
Amino Acid Catabolism & Synthesis - Protein Turnover
Although essential amino acids are not stored as such in the body for any significant period of time, all body proteins are continuously being broken down and resynthesized in a process known as protein turnover.¹² ¹⁵ ¹⁷ During protein turnover, some amino acids enter catabolic pathways and are permanently lost. Therefore, adequate amounts of high-quality dietary protein, containing all the essential amino acids, must be consumed each day to replace both the essential and nonessential amino acids lost to catabolism.
Hypermetabolic states, such as hyperthyroidism or other illness, increase both protein turnover and nitrogen losses and therefore increase the daily protein requirements.¹⁸ ¹⁹
Protein as an Energy Source
Unlike fat or carbohydrate, protein cannot be stored as such in the body, other than as muscle protein itself. In animals fed diets containing more protein than is needed, the extra protein is metabolized and used for energy. See Figures 1 and 2.
In all species, but most importantly in cats, ingested amino acids are converted to carbohydrates via gluconeogenesis.¹² ¹⁵ ¹⁷ ²⁰⁻²² This pathway is also used under starvation conditions to generate glucose and energy from the body's own proteins, particularly those found in muscle.
Factors That Determine How Dietary Proteins are Used
- All or None Rule: To make a certain protein, the necessary amino acids must be present in the cell in the right amounts, or the protein will not be made. Essential amino acids that are not used to make proteins are not stored.
- Adequacy of Caloric Intake: If the diet fails to provide sufficient calories as carbohydrates and fats, proteins will not be synthesized; instead the ingested amino acids will be used as a source of energy.
- Nitrogen Balance: Normally, the amount of protein synthesis is equal to the amount of protein breakdown.¹² ¹⁷ If there is more protein synthesis than breakdown, then we have a positive nitrogen balance, e.g., recovery from injury. If there is greater protein breakdown than synthesis, then there is a negative nitrogen balance, e.g, starvation, illness, hyperthyroidism.
- Hormonal Controls: Anabolic hormones, e.g., insulin, growth hormone, sex hormones, stimulate the production or maintenance of proteins. Catabolic hormones, e.g, glucocorticoids, thyroid hormone, stimulate the breakdown of proteins.¹⁸ ¹⁹
Cats Do Not Need Carbohydrate, But Have High Protein Requirements
 The cat, as a strict carnivore, has evolved to depend on protein as a major energy source. The natural diet of cats in the wild is based upon the consumption of small mammals, birds and insects - the composition of this diet is high in protein and fat but low in carbohydrate.²⁰⁻²² This natural diet high in animal protein contains all of a cat's essential amino acids, which is not the case for plant-based proteins.
The cat, as a strict carnivore, has evolved to depend on protein as a major energy source. The natural diet of cats in the wild is based upon the consumption of small mammals, birds and insects - the composition of this diet is high in protein and fat but low in carbohydrate.²⁰⁻²² This natural diet high in animal protein contains all of a cat's essential amino acids, which is not the case for plant-based proteins.
Cats have no dietary requirement or need for carbohydrate but are adapted physiologically and metabolically for high protein intake.²³ In support of that, normal cats do well when fed diets containing 70 percent protein.²⁴⁻²⁷ The lack of a need for dietary carbohydrates is related to the fact that cats have developed a tremendous ability to synthesize their needed glucose from protein catabolism via hepatic gluconeogenesis.²⁴ ²⁵ ²⁸
Cats have a much higher protein requirement than other species, such as dogs and humans. The protein requirement for the adult cat is 2-3 times as much as the adult dog, and their protein requirement increases even further as cats reach old age.³ ⁸⁻¹¹ ²³ This high protein requirement of cats is primarily related to the fact that cats have a limited ability to decrease the hepatic enzymes responsible for amino acid catabolism, even when fed a lower than optimal protein intake.²⁴ ²⁹ ³⁰
The Domestic Cat: A Metabolically Inflexible Carnivore
When most omnivores, e.g., humans, rats, pigs, dogs, ingest a diet high in protein, the activities of the amino acid-catabolizing enzymes in the liver increase to cope with the higher flux of amino acids.³¹⁻³³ The activity of the urea cycle enzymes also increase to metabolize the increased ammonia generated upon the catabolism of the amino acids. On the other hand, when fed a diet low in protein, omnivores accommodate by lowering the hepatic activity of these catabolic enzymes of amino acid metabolism, as well as decreasing hepatic urea production.³¹⁻³³
By contrast, cats have a very limited ability to down-regulate these hepatic catabolic enzymes when fed a low-protein diet. In a classic study, Rogers, et al, compared the activity of several catabolic enzymes of amino acid metabolism in adult cats fed either a high- or low-protein diet or fasted for five days.29 Results showed little changes in the hepatic enzyme activities between the three groups of cats, with hepatic enzyme activities remaining set at high levels to cope with a high protein diet, even when they weren't being fed!
Why would cats be so different? Well, compared to other carnivores, cats may not be so strange after all. A similar degree of metabolic "inflexibility" has also been reported in other carnivores such as barn owls and the rainbow trout.³⁴ ³⁵
This limited metabolic flexibility in cats and other obligate carnivores likely represents an evolutionary adaptation to a consistently abundant supply of dietary protein. The moderately high and fixed activity of the urea cycle provides a safeguard against ammonia toxicity after ingestion of a high-protein meal. In addition, the high rate of amino acid catabolism allows for a readily available source of energy via direct oxidation or as a substrate for hepatic gluconeogenesis.
It is only when a cat is fed a lower protein diet - a condition that would never happen in the wild - that the high rate of protein catabolism becomes a disadvantage.
Additional Reading
Care to Compare? Wild vs. Domesticated Prey
Answers: Why Did My Cat's Fur Get So Silky?
Bottom Line
- Unlike omnivores such as dogs, pigs, rats or humans, cats have a limited ability to decrease the activity of the hepatic enzymes responsible for removing amino groups from the amino acids when fed a low protein diet.
- Because these feline hepatic enzyme systems are constantly active, a fixed amount of dietary, or muscle, protein will always be catabolized for energy no matter how much energy in the form of carbohydrate or fat the cat ingests.
- In addition, neither fat nor carbohydrates can be directly converted to amino acids to be made into protein. In this regard, the carnivorous cat is similar to omnivores.
- Overall, this explains why muscle wasting can occur so quickly in the older, geriatric cat, which becomes ill or develops a poor appetite or is fed a low-protein diet.
Dr Mark E. Peterson has been deeply involved in clinical research for over 35 years, and remains at the forefront of the science that advances the study and knowledge of endocrine diseases of cats. He was the first veterinarian to document hyperthyroidism in cats (1979) and the first to treat hyperthyroid cats with radioiodine (in 1980). In addition to hyperthyroidism, Dr. Peterson was the first to document a number of "new" diseases in cats, including acromegaly, hypoparathyroidism, insulinoma and Addison's disease. Dr. Peterson is director of theAnimal Endocrine Clinicin New York City, a specialty referral hospital devoted exclusively to the care of cats and dogs with endocrine disease.
1. G Perez-Camargo, "Cat nutrition: What is new in the old?" Compendium for Continuing Education for the Practicing Veterinarian 26, Suppl 2A, 5-10.
2. D Laflamme, "Nutrition for Aging Cats and Dogs and the Importance of Body Condition," Veterinary Clinics of North America: Small Animal Practice 35, 2005, 713-742.
3. G Pérez-Camargo, "Feline Decline in Key Physiological Reserves: Implication for Mortality," Proceedings of the Nestlé Purina Companion Animal Nutrition Summit: Focus on Gerontology, St. Louis, MO, 2010, 6-13.
4. S Fujita and E Volpi, "Nutrition and Sarcopenia of Ageing," Nutrition Research Reviews 17, 2004, 69-76.
5. D Paddon-Jones, KR Short, WW Campbell, et al, "Role of Dietary Protein in the Sarcopenia of Aging," The American Journal of Clinical Nutrition 87, 2008, 1562S-1566S.
6. KR Short and KS Nair, "Mechanisms of Sarcopenia of Aging," Journal of Endocrinological Investigation 22, Suppl 5, 1999, 95-105.
7. RR Wolfe, "Sarcopenia of Aging: Implications of the Age-related Loss of Lean Body Mass," Proceedings of the Nestlé Purina Companion Animal Nutrition Summit: Focus on Gerontology, St. Louis, MO, 2010, 12-17.
8. SE Little, "Evaluation of the Senior Cat with Weight Loss," In: SE Little, (ed), The Cat: Clinical Medicine and Management, Philadelphia, Elsevier Saunders, 2012, 1176-1181.
9. AH Sparkes, "Feeding Old Cats - An Update on New Nutritional Therapies," Topics in Companion Animal Medicine, no. 26, 2011, 37-42.
10. AR Patil, C Cupp and G Pérez-Camargo, "Incidence of Impaired Nutrient Digestibility in Aging Cats," Nestlé Purina Nutrition Forum Proceedings 26, 2003, Suppl 2A, 72.
11. JJ Wakshlag, "Dietary Protein Consumption in the Healthy Aging Companion Animal," Proceedings of the Nestlé Purina Companion Animal Nutrition Summit: Focus on Gerontology, St. Louis, MO, 2010, 32-39.
12. KL Gross, RM Yamka, C Khoo, et al, "Macronutrients," In: MS Hand, CD Thatcher, RL Remillard, R Roudebush, BJ Novotny, (eds), Small Animal Clinical Nutrition, Mark Morris Institute, 2010, 49-105.
13. KP Davy, T Horton, BM Davy, et al, "Regulation of Macronutrient Balance in Healthy Young and Older Men," International Journal of Obesity and Related Metabolic Disorders 25, 2001, 1497-1502.
14. J Galgani and E Ravussin, "Energy Metabolism, Fuel Selection and Body Weight Regulation," International Journal of Obesity 32, Suppl 7, 2008, S109-119.
15. T Brody, Nutritional Biochemistry. Second Edition, Academic Press. 1998.
16. New Oxford Dictionary of English, Oxford University Press, 2001.
17. JM Berg, JL Tymoczko and L Stryer, "Protein Turnover and Amino Acid Catabolism," Biochemistry, 5th edition, New York, WH Freeman, 2002.
18. WE Mitch, "Mechanisms Accelerating Muscle Atrophy in Catabolic Diseases," Transactions of the American Clinical and Climatological Association 111, 2000, 258-269.
19. AL Riis, JO Jørgensen, S Gjedde, et al, "Whole Body and Forearm Substrate Metabolism in Hyperthyroidism: Evidence of Increased Basal Muscle Protein Breakdown," American Journal of Physiology: Endocrinology and Metabolism 288E, 2005, 1067-1073.
20. A Myrcha and J Pinowski, "Weights, Body Composition and Caloric Value of Post-juvenile Molting European Tree Sparrows," Condor, no. 72, 1970, 175-178.
21. JF Vondruska, "The Effect of a Rat Carcass Diet on the Urinary pH of the Cat," Companion Animal Practice, no. 1, 1987, 5-9.
22. SD Crissey, KA Slifka and BA Lintzenich, "Whole Body Cholesterol, Fat and Fatty Acid Concentrations of Mice (Mus domesticus) Used as a Food Source," Journal of Zoo and Wildlife Medicine, no. 30, 1999, 222-227.
23. National Research Council, "Proteins and Amino Acids," In: Nutrient Requirements of Dogs and Cats, Washington, DC: National Academies Press, 2006, 111-145.
24. ML MacDonald, QR Rogers and JG Morris, "Nutrition of the Domestic Cat, A Mammalian Carnivore," Annual Review of Nutrition, no. 4, 1984, 521-562.
25. JG Morris, "Idiosyncratic Nutrient Requirements of Cats Appear to be Diet-induced Evolutionary Adaptations," Nutrition Research Reviews, no. 15, 2002, 153-168.
26. WH Hendriks, "Protein Metabolism in the Adult Domestic Cat (Felis catus)," PhD Thesis, 1996.
27. B Hamper, J Bartges, C Kirk, et al, "The Unique Nutritional Requirements of the Cat: A Strict Carnivore," In: SE Little, ed., The Cat: Clinical Medicine and Management, St. Louis: Elsevier Saunders, 2012, 236-242.
28. IC Kettelhut, MC Foss and RH Migliorini, "Glucose Homeostasis in a Carnivorous Animal (Cat) and in Rats Fed a High-protein Diet," American Journal of Physiology 239, 1980, R437-440.
29. QR Rogers, JG Morris and RA Freedland, "Lack of Hepatic Enzymatic Adaptation to Low and High Levels of Dietary Protein in the Adult Cat," Enzyme 22, 1977, 348-356.
30. AS Green, JJ Ramsey, C Villaverde, et al., "Cats are Able to Adapt Protein Oxidation to Protein Intake Provided Their Requirement for Dietary Protein is Met," The Journal of Nutrition, 2008, 1053-1060.
31. JC Waterlow JC, "The Mysteries of Nitrogen Balance," Nutrition Research Reviews 12, 1999, 25-54.
32. RW Rosebrough, NC Steele and JP McMurtry, "Effect of Protein Level and Supplemental Lysine on Growth and Urea Cycle Enzyme Activity in the Pig," Growth 47, 1983, 348-260.
33. RT Schimke, "Adaptive Characteristics of Urea Cycle Enzymes in the Rat," Journal of Biological Chemistry 237, 1962, 459-68.
34. ML Walton, "Metabolic Effects of Feeding a High Protein/low Carbohydrate Diet as Compared to a Low Protein/high Carbohydrate Diet to Rainbow Trout Salmo Gairdneri," Fish Physiology and Biochemisty 1, 1986, 7-15.
35. MR Meyers and KC Klasing, "Low Glucokinase Activity and High Rates of Gluconeogenesis Contribute to Hyperglycemia in Barn Owls (Tyto alba) After a Glucose Challenge," Journal of Nutrition 129, 1999, 1896-1904.
